|
大隅グループ adult brain チームでは、生後脳におけるニューロン新生の分子機構と精神機能の関わりについて注目し、主に以下のようなテーマで研究しています。
 生後脳の海馬における神経新生の分子メカニズム 生後脳の海馬における神経新生の分子メカニズム
大脳皮質におけるグリア細胞分化の分子メカニズム
Pax6 ヘテロ変異体の行動解析
嗅球介在ニューロン分化のメカニズム
生後脳の海馬における神経新生の分子メカニズム
近年、成体においてニューロンは側脳室(SVZ) および海馬歯状回(SGZ)において新生することが再発見され、現在とくに脳の高次機能との関連性が指摘されています。
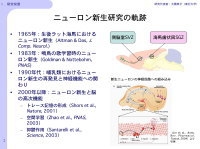
【図1】成体における神経新生研究の軌跡
私たちの研究室では、転写制御因子Pax6の胎児における機能解析を行ってきましたが、Pax6が生後の脳においても発現していることを見出し、とくに海馬の歯状回とよばれる領域にPax6陽性細胞が存在することから
(図2)、Pax6ヘテロ変異ラットにおいて海馬の神経新生について解析しました(Maekawa et al., 2005)。
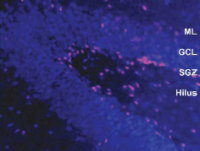
【図2】成体海馬におけるPax6陽性細胞の分布
野生型ラットにおいてPax6はSGZの幹細胞様の細胞に発現している。
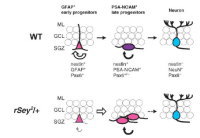
興味深いことに、Pax6は神経前駆細胞に発現し、変異ラット海馬においては、BrdUを取り込む細胞の数が野生型に比べ減少し、マーカー解析から神経幹細胞の性質が変化していることが明らかになりました (Maekawa et al., 2005)。このような解析から、Pax6は神経幹細胞の維持に寄与していると考えています(図3)。
またニューロン新生と精神機能において幾つかの関連性が示されており(図4)、脳の発達と環境因子に関して、不飽和脂肪酸との関連性についても検討する予定です。
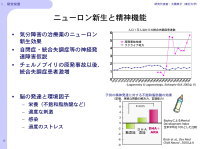
【図4】神経新生と精神疾患との関わり
現在までに神経新生に関わる因子が多く同定されてきていますが、私たちの目標として、Pax6変異ラットを材料にして、Pax6の下流の遺伝子カスケードを理解したいと考えています(図5)。ニューロン新生におけるPax6の役割を詳細に解析するため、マイクロアレイを用いたPax6下流因子の網羅的解析も行う予定であり、胎生期でPax6の下流遺伝子として同定されているいくつかの因子についても、成体における神経新生への関与があるかどうかを検討しています。
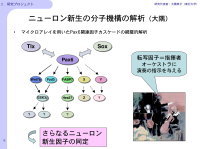
【図5】神経新生と精神疾患との関わり
大脳皮質におけるグリア細胞分化の分子メカニズム
高次の脳機能は神経細胞のみの機能によって成立するものではなく、それらを取り巻くグリア細胞の存在も重要です。Pax6はグリア系の細胞にも発現しており、これらの細胞分化におけるPax6の役割についても解析しています(図6)。また、Pax6のさらなる機能解析のため、条件つき遺伝子破壊法を行う予定です。
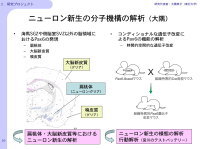
【図6】成体におけるPax6の発現領域と細胞種
Pax6 ヘテロ変異体の行動解析
三菱化学生命科学研究所の井ノ口先生のグループと共同研究でPax6変異ラット(成体ヘテロ)の行動解析を行っています。Pax6変異ラットでは恐怖心の低下やプレパルス抑制 (図7) の低下(統合失調症の指標)などが認められます(未発表)。このことから、Pax6変異ラットはヒト統合失調症のモデル動物としても利用できそうです(特許出願中)。また今後、脳の発生発達過程におけるPax6の機能について詳細に解析する予定です。ニューロン新生と精神機能において幾つかの関連性が示されています。

【図7】プレパルス抑制について
嗅球介在ニューロン分化のメカニズム
成体では海馬の他に、側脳室の細胞からニューロンが産生され、嗅球の介在ニューロンを作ることが知られています。Pax6は移動中の細胞に発現することが知られており、Pax6変異マウスでは嗅球がやや小さくなる表現型が見られます。そこで、現在、嗅球介在ニューロンの分化におけるPax6の役割について解析を進めています。
|