若松グループ
神経堤細胞
神経堤細胞は脊椎動物にユニークな細胞集団で、発生過程において一過的に現れます。胚の体内を広範囲に移動し(図1)、末梢神経系のニューロンやグリア、色素細胞、顎等の頭部組織、大脳の血管系を構成する周辺細胞など、驚くほど多様な細胞種に分化します。したがって、神経堤症と総称される遺伝病では神経堤細胞の発生に異常が生じることで様々な組織に問題が起きてしまいます。
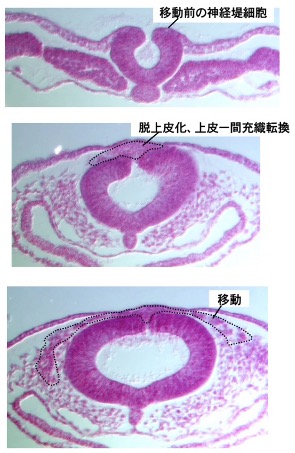
図1:神経堤の初期発生。ニワトリ胚の切片画像。神経管の背側領域に誘導された神経堤細胞(上段)は、上皮—間充織転換を経て(中段)、胚体内を移動しはじめる。
神経堤細胞の発生を制御する遺伝子たち
当研究グループでは、神経堤細胞の発生がどのように制御されているのかを明らかにしてきました。例えば神経堤細胞の分化制御については、Notchシグナルの非対称性がニューロンvs幹細胞の運命決定をおこなっていること(Wakamatsu et al., 2000)や、転写因子のSox2がニューロン分化を抑制すること(Wakamatsu et al., 2004, Wakamatsu and Uchikawa, 2021)等を示してきました。また、神経堤は最初上皮性の外胚葉組織に誘導されますが、Notchシグナルを介して活性化されるBMP4が神経堤細胞の誘導に必須であることを示しました(Endo et al., 2002, 2003)。さらに、誘導された神経堤細胞は上皮−間充織転換を経て移動能力を獲得しますが(図1参照)、この際にPKAシグナルやSox9、ATF4といった転写制御因子が働いていることを明らかにしています(Sakai et al., 2005, 2006, Suzuki et al., 2006, 2010, Wakamatsu and Uchikawa, 2021)。
頭部神経堤細胞の発生と進化
頭部神経堤細胞からは顎などの頭部組織が作られます。逆に、神経堤細胞が脊椎動物の進化の過程で獲得され、はじめて頭部というものができたと言えます。有袋類(袋を持つ哺乳類、カンガルーやコアラを想像してください)では、この頭部神経堤細胞の発生が非常に早くおきます(図2)。これは、有袋類が子供を未熟な状態で産み、それを母乳で育てるという生態をもっているためです。なにしろ、顎ができていないと母乳が飲めませんから。
このような発生様式の時間的変更のことを「ヘテロクロニー(異時性)」といいますが、私たちはこの有袋類における頭部神経堤細胞のヘテロクロニーがどのようなメカニズムで制御されているのか、小型有袋類のオポッサムを用いて明らかにしようと試みています。実験系として確立されたものが少ないので、チャレンジングなプロジェクトになっています。これまでに、神経堤の形成に重要であるSox9遺伝子がオポッサム胚ではマウスやニワトリと比べて非常に早い発生段階で予定神経堤領域に発現すること(Wakamatsu et al., 2014)、有袋類のゲノムではSox9遺伝子の神経堤エンハンサーに特有の配列が含まれており、これがSox9遺伝子の発現を早めること(Wakamatsu & Suzuki, 2019)などを明らかにしてきました。
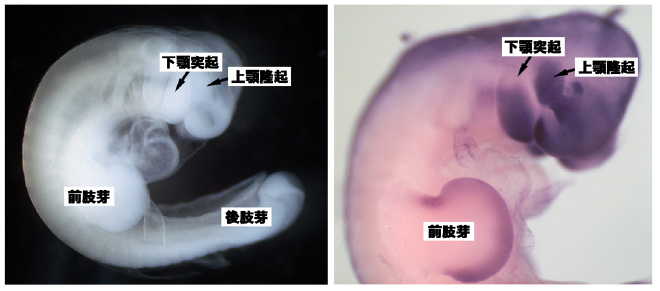
図2:オポッサムの11.5日胚(左)と、Msx1遺伝子の発現(右)。11.5日胚のオポッサム胚では、顎原基がすでに大きく発達している。また、産道から乳首まで這っていくために必要な前肢も、後肢より発達してきている。Msx1遺伝子はこれらの組織で強く発現している(紫の染色)。
顎の発生と哺乳類における異形歯性の進化
顎の原基は神経堤に由来する間充織とそれを覆う上皮から作られます。顎の原基では基部から先端部にかけて異なるホメオボックス遺伝子が発現しており、その組み合わせが顎の形態を制御しています。哺乳類では顎の基部–先端部軸にそって異なる形態/機能の歯(先端から切歯、犬歯、小臼歯、臼歯の順)が生えますが、これを「異形歯性」と呼びます。そして、顎原基の神経堤由来間充織細胞がどのホメオボックス遺伝子を発現するかによって、そこから生えてくる歯のタイプを決定すると提唱されています(ホメオボックスコード仮説、McCollum and Sharpe, 2001)
私たちは、全ての歯のタイプを持つオポッサムやフェレットの胚と、切歯と臼歯しかもたないマウスの胚の顎原基についてホメオボックス遺伝子の発現を比較しました(Msx1の発現は図2参照)。この研究によって、ホメオボックス遺伝子群のうちAlx3+Msx1の組み合わせが切歯、Msx1のみが犬歯、Msx1+BarX1の組み合わせが小臼歯、BarX1のみが臼歯と対応していること(図3)、マウスの下顎ではMsx1とBarX1両方が発現する領域が極端に狭くなっており、これがマウスを含むげっ歯類における歯のパターンを決定している可能性を示しました(Wakamatsu et al., 2019)。Msx1遺伝子の発現の広さが重要であることが示唆されたことから、顎原基におけるMsx1遺伝子の発現制御について解析を進めています。ヒトにおいてMsx1遺伝子の先天異常は一部の歯の欠損の原因になることも知られており、本研究は哺乳類の進化だけでなく、医学的にも重要な意義を持ちます。
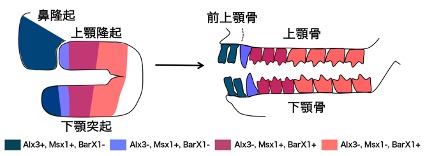
図3:オポッサム胚の顎原基におけるホメオボックス遺伝子群の発現領域(左)と、それに対応した歯のタイプ(右)。Alx3、Msx1、BarX1の発現の組み合わせによって切歯、犬歯、小臼歯、臼歯の生える場所が規定される。
哺乳類の脳の進化
上記の神経堤/神経堤由来組織の発生と進化についての他に、当研究室の吉川らとの共同研究として、哺乳類の脳がどのように進化してきたかについても研究を行なっています。